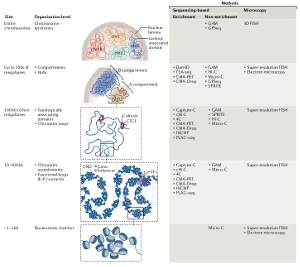
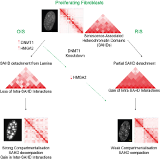
The genome is more than a linear string of genes. It has secondary, tertiary and quaternary higher-order structures. Strings of genes organize in chromosomal domains that are characterized by one of relatively few different types of chromatin, either repressive or activating. In the 3D space, each of the elements in one domain makes frequent contacts with other elements in the same domain. These entities have been called physical- or topologically associated- domains (TADs).
At a chromosome level, individual TADs form contacts with other TADs, preferentially of the same type, in order to build ordered 3D architectures that are called chromosome territories. Finally, different chromosomes organize non randomly in the nuclear space.
Therefore, the eukaryotic genome is highly organized in 3D and this regulation can be transmitted or modulated during the life of cells and organisms.
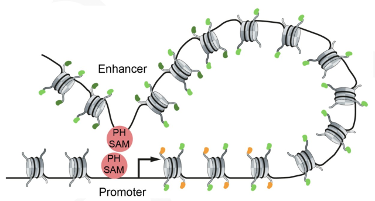
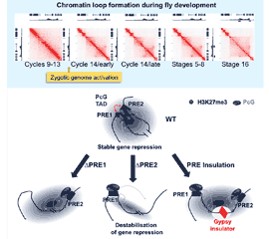
This sophisticated chromosomal choreography involves thousands of different players – DNA sequences, RNAs and proteins – but rather than combining in infinite numbers of ways, these components organize a relatively limited number of types of chromatin, either active or repressive. In particular, two main groups of genome regulatory components are proteins of the Polycomb Group (PcG) and of the trithorax Group (trxG). PcG proteins are maintain the memory of silent states of gene expression through cell physiology and multiple cell divisions, while trxG members maintain active chromatin states
These proteins are able to recognize regulatory states of their target genes and to maintain these states even after disappearance of the primary transcriptional regulators that have induced them in the first place. Remarkably, these states can also be transmitted to a fraction of the progeny over multiple generations. In our lab, we aim at understanding the principle governing 3D genome organization, its functional implications, and the molecular mechanisms by which PcG and trxG proteins regulate their target genes, convey inheritance of chromatin states and orchestrate development.
To reach this goal, we employ a variety of complementary approaches and techniques in the areas of molecular, cellular and developmental biology, genomics and bioinformatics.
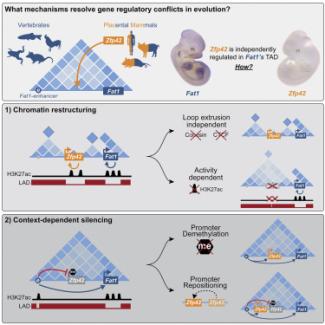
Repression and 3D-restructuring resolves regulatory conflicts in evolutionarily rearranged genomes.
Ringel AR, Szabo Q, Chiariello AM, Chudzik K, Schöpflin R, Rothe P, Mattei AL, Zehnder T, Harnett D, Laupert V, Bianco S, Hetzel S, Glaser J, Phan MHQ, Schindler M, Ibrahim DM, Paliou C, Esposito A, Prada-Medina CA, Haas SA, Giere P, Vingron M, Wittler L, Meissner A, Nicodemi M, Cavalli G, Bantignies F, Mundlos S, Robson MI
A shared ancient enhancer element differentially regulates the bric-a-brac tandem gene duplicates in the developing Drosophila leg.
Bourbon HG, Benetah MH, Guillou E, Mojica-Vazquez LH, Baanannou A, Bernat-Fabre S, Loubiere V, Bantignies F, Cavalli G, Boube M
Comprehensive characterization of the epigenetic landscape in Multiple Myeloma.
Alaterre E, Ovejero S, Herviou L, de Boussac H, Papadopoulos G, Kulis M, Boireau S, Robert N, Requirand G, Bruyer A, Cartron G, Vincent L, Martinez AM, Martin-Subero JI, Cavalli G, Moreaux J
Challenges and guidelines toward 4D nucleome data and model standards.
Marti-Renom MA, Almouzni G, Bickmore WA, Bystricky K, Cavalli G, Fraser P, Gasser SM, Giorgetti L, Heard E, Nicodemi M, Nollmann M, Orozco M, Pombo A, Torres-Padilla ME
Loss of PRC1 induces higher-order opening of Hox loci independently of transcription during Drosophila embryogenesis.